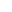如何选择限制性内切酶?酶切位点到底应该怎么设计,才能提高克隆成功率?
对于刚接触分子克隆的新手来说,设计表达载体时往往会遇到这样的问题:
这些问题看似都发生在实验台上,实际上很多在设计阶段就已经埋下了隐患。
限制性内切酶(Restriction Enzymes)不仅是 DNA 克隆中最常用的工具之一,酶切位点的选择更直接决定了后续酶切、连接、转化乃至蛋白表达是否能够顺利进行。

传统限制性内切酶克隆法介导的质粒间DNA片段亚克隆简易流程示意图
本文将围绕实际实验中的常见问题,介绍如何选择合适的限制性内切酶,并讲清楚酶切位点设计中最容易忽视的关键细节。
限制性内切酶是一类能够识别特定 DNA 序列,并在特定位点切开双链 DNA 的酶。
例如,最经典的 EcoRI 识别序列为:5'-GAATTC-3'
当 DNA 中出现这段序列时,EcoRI 就会识别并切开,形成带有黏性末端(Sticky End)的 DNA 片段。
不同限制性内切酶具有不同的识别序列。例如:
| 限制性内切酶 | 识别序列 | 末端类型 |
|---|---|---|
| EcoRI | GAATTC | 黏性末端 |
| XhoI | CTCGAG | 黏性末端 |
| BamHI | GGATCC | 黏性末端 |
| HindIII | AAGCTT | 黏性末端 |
| NotI | GCGGCCGC | 黏性末端 |
| EcoRV | GATATC | 平末端(Blunt End) |
这些酶共同构成了分子克隆中最基础的"DNA 剪刀"。
拿到一个表达载体后,很多新手第一反应就是找一个熟悉的酶切位点。
实际上,正确的第一步应该是查看多克隆位点(MCS)。

MCS 是载体中人工设计的一段短 DNA 序列,集中包含多个唯一的限制性内切酶识别位点,方便研究者插入外源基因。
例如,一个典型的 MCS 可能包含:
这些位点通常各出现一次,因此非常适合用于克隆。
设计时应优先选择位于 MCS 中、且在整个载体中仅出现一次的酶切位点,以避免切开载体其他区域。
假设你的目的基因内部本身就含有一个 EcoRI 识别序列:GAATTC
如果仍然使用 EcoRI 对 PCR 产物进行酶切,那么目的基因也会被切成两段。
最终不仅无法连接,还可能得到完全错误的克隆结果。因此,在设计之前,一定要检查目的基因内部是否包含所选限制性内切酶的识别序列。
目前常用的软件包括:
这些软件都可以自动显示载体和插入片段中的限制性内切酶位点,帮助研究者快速筛选合适的酶。

初学者有一个常见疑问:既然 EcoRI 可以切开载体,为什么不能两端都用 EcoRI?原因在于方向性(Directionality)。
如果载体和插入片段两端完全相同,那么连接时可能出现正向插入、反向插入、载体自连等多种情况,其中正确方向的克隆往往占比很低,需要大量筛选。
如果目的基因方向反了,即使成功连接,也可能无法正确表达。因此,在表达载体构建中,更推荐使用两种不同的限制性内切酶,例如:
5' EcoRI ------ Gene ------ XhoI 3'
这样两端产生不同的黏性末端,只能按一个方向连接,大大提高正确克隆的比例。这也是所谓的定向克隆(Directional Cloning)。
限制性内切酶切割 DNA 后,可以产生两种类型的末端:
黏性末端具有互补的单链突出端,能够先通过碱基配对暂时结合,再由 DNA 连接酶完成连接,因此:

相比之下,平末端没有互补突出端,DNA 完全依赖连接酶随机连接,因此:

因此,在常规表达载体构建中,除非实验设计有特殊要求,大多数研究者都会优先选择能够产生黏性末端的限制性内切酶。
成功完成克隆,并不意味着蛋白一定能够正确表达。如果载体带有 His、FLAG、HA、Myc 或 GFP 等标签,还必须确认插入片段与标签保持相同的阅读框。
例如,一个碱基的插入或缺失,就可能导致整个开放阅读框发生移码(Frameshift)。
其结果可能包括:
因此,在提交引物合成前,建议使用 SnapGene 或 Benchling 模拟克隆结果,检查起始密码子(ATG)、标签以及终止密码子是否位于同一阅读框。

很多双酶切实验失败,并不是因为 DNA 有问题,而是因为两种酶的反应条件并不兼容。
不同限制性内切酶对缓冲液组成、盐浓度和反应温度的要求可能不同。如果两种酶不能在同一体系中保持较高活性,就需要分步酶切,增加实验时间,也可能导致 DNA 损失。
因此,在选择双酶切组合时,应优先考虑:
许多厂商会在产品说明中提供兼容性信息,也可以利用在线工具快速查询酶切条件。
限制性内切酶和酶切位点设计,是分子克隆中最基础却也最容易被忽视的一环。一个合理的酶切方案,不仅能够提高克隆效率,还能减少空载体、自连、方向错误和移码等问题,为后续蛋白表达和功能研究奠定基础。
随着无缝克隆(Gibson Assembly、In-Fusion、Golden Gate Assembly 等)等新技术的发展,传统限制性内切酶克隆虽然不再是唯一选择,但由于其成本低、操作成熟、适用范围广,仍然是生命科学实验室中最经典、最常用的分子克隆方法之一。